












HAP1基因编辑细胞模型:单倍体优势赋能与精准疾病模型平台

HAP1细胞系源自慢性髓系白血病患者KBM-7细胞,经化学诱变后获得了近乎单倍体的核型。虽然后续染色体分析表明其部分区域可能已变为二倍体,但整体基因拷贝数已简化,因此被广泛用作“类单倍体”的人类遗传筛选工具。
这种近乎单倍体的特性意味着大多数基因只有一个拷贝,一旦用CRISPR进行编辑,基因敲除的表型会立刻显现,从而避免了二倍体细胞中因另一等位基因补偿带来的干扰。
HAP1已成为功能性遗传研究中“最受欢迎的细胞模型之一”。早在2015年,《Nature Reviews Genetics》的研究亮点就曾聚焦于利用HAP1进行的全基因组筛选——The genetic essence of human cells一文重点报道了两项同期发表在Science杂志上的独立研究。这两项研究均利用单倍体细胞系(包括HAP1)进行全基因组筛选,鉴定了人类细胞的“核心必需基因组”。
01
为什么HAP1是科研领域的“利器”?
1. 单倍体优势:让基因编辑更直接
HAP1细胞最核心的特点在于其近乎单倍体的核型,基因组结构相对简单,大多数基因在基因组中只有一个功能拷贝。这意味着利用CRISPR/Cas9技术进行基因敲除时,编辑效率更高,表型鉴定更直接,大大缩短了实验周期。
2. 贴壁生长与操作的便利性
与悬浮细胞相比,HAP1细胞呈贴壁生长,形态均一,便于进行各类生化分析和显微成像。其倍增时间短,易于扩增,非常适合大规模筛选实验。
3. 大规模多组学研究验证HAP1的强大潜力
2022年,Rensvold等人在Nature上发表了一项里程碑式研究。他们利用CRISPR技术在HAP1细胞中构建了203个线粒体基因敲除细胞系,并通过蛋白质组、脂质组、代谢组等多组学分析,系统解析了数百个线粒体蛋白的功能,发现了PYURF(NDUFAFQ)、SLC30A9、RAB5IF等新功能基因,并为未确诊的线粒体病患者提供了分子诊断依据。这一研究充分证明了HAP1敲除细胞在大规模、系统性功能基因组学研究中的巨大潜力。
02
基因编辑HAP1细胞的关键应用领域
基因编辑HAP1细胞系的应用领域极其广泛,涵盖了从基础细胞生物学到转化医学的各个层面。以下结合部分代表性基因,阐述其在关键研究领域中的应用。
1. 激酶与信号转导:从靶点发现到耐药机制研究
激酶是细胞信号网络的核心调控者,其功能异常与肿瘤、免疫疾病及代谢紊乱密切相关。HAP1敲除细胞为解析激酶的生物学功能和验证其作为药物靶点提供了理想平台。
● PI3K-AKT信号通路与代谢重编程:
PTEN是PI3K-AKT信号通路的负调控因子,其缺失是多种癌症中的常见事件。PTEN缺失导致mTORC1过度激活是肿瘤生物学领域的经典发现。
利用PTEN基因敲除HAP1细胞,研究者能够清晰地复现PTEN缺失后mTORC1信号通路的过度激活,并深入探究其对细胞增殖、代谢重编程和自噬的调控机制。该细胞模型为筛选靶向PTEN缺失肿瘤的mTOR抑制剂、理解肿瘤代谢脆弱性提供了关键的实验工具。
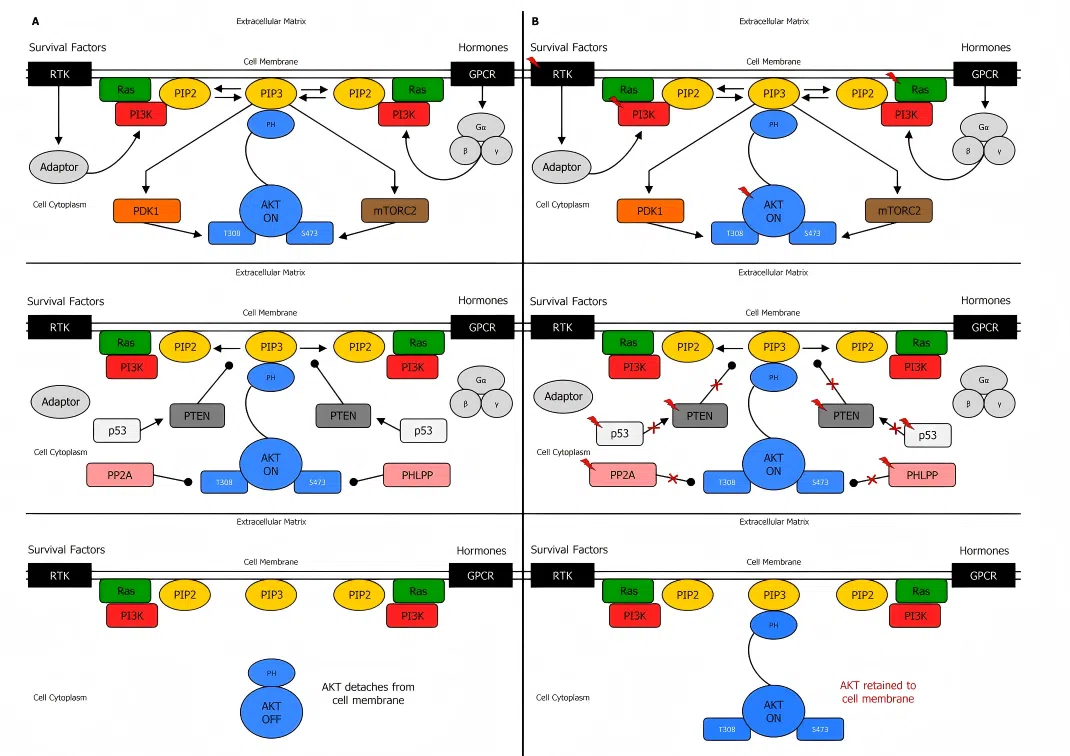 图1. PI3K、PTEN 和 AKT 调控的生化机制
● 免疫与肿瘤微环境:
图1. PI3K、PTEN 和 AKT 调控的生化机制
● 免疫与肿瘤微环境:
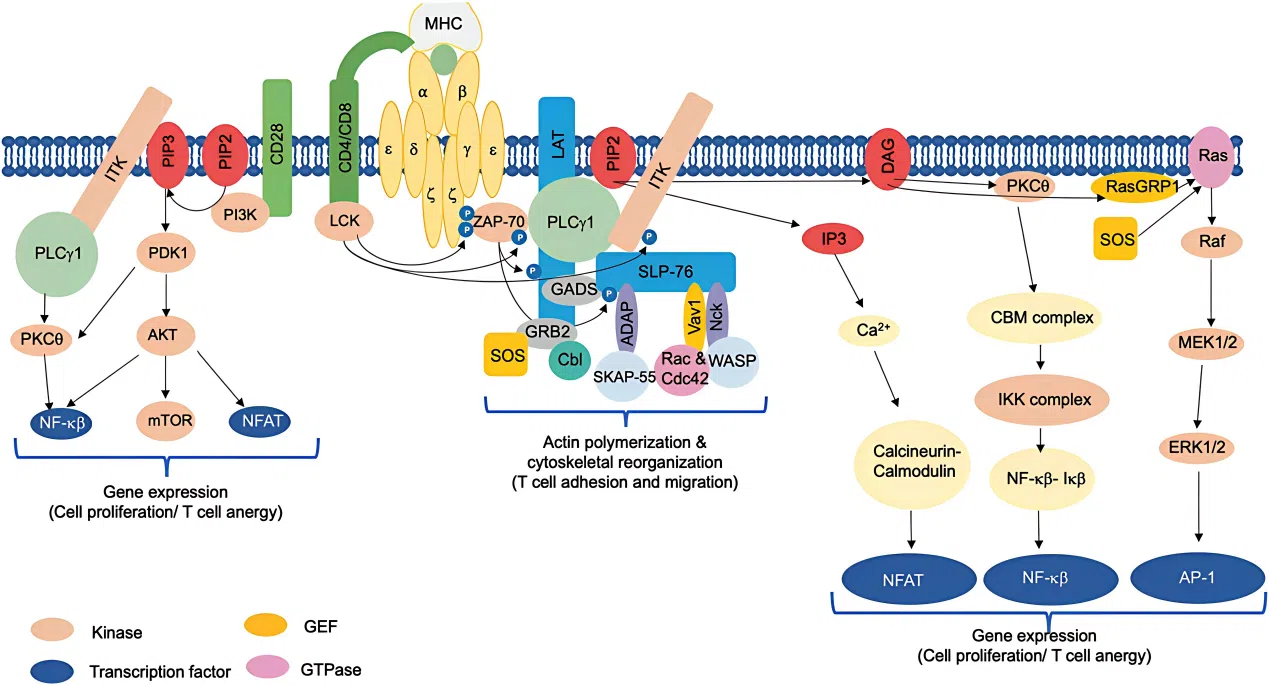
蛋白激酶Cθ(PKCθ,由PRKCQ基因编码)属于丝氨酸/苏氨酸蛋白激酶家族,其激活不依赖钙离子,但需要磷脂作为辅助因子。该激酶在T细胞受体介导的信号通路中发挥核心调控作用,是T细胞活化所必需的关键分子。由于其在免疫应答中的独特地位,PKCθ已成为自身免疫性疾病和肿瘤免疫治疗领域的重要靶点。
利用PRKCQ基因敲除的HAP1细胞,可以在同基因背景下精准评估该激酶功能缺失对T细胞信号转导的影响,为靶向PKCθ的药物筛选和机制研究提供可靠的细胞模型。
PTEN是PI3K-AKT信号通路的负调控因子,其缺失是多种癌症中的常见事件。PTEN缺失导致mTORC1过度激活是肿瘤生物学领域的经典发现。
利用PTEN基因敲除HAP1细胞,研究者能够清晰地复现PTEN缺失后mTORC1信号通路的过度激活,并深入探究其对细胞增殖、代谢重编程和自噬的调控机制。该细胞模型为筛选靶向PTEN缺失肿瘤的mTOR抑制剂、理解肿瘤代谢脆弱性提供了关键的实验工具。
图1. PI3K、PTEN 和 AKT 调控的生化机制蛋白激酶Cθ(PKCθ,由PRKCQ基因编码)属于丝氨酸/苏氨酸蛋白激酶家族,其激活不依赖钙离子,但需要磷脂作为辅助因子。该激酶在T细胞受体介导的信号通路中发挥核心调控作用,是T细胞活化所必需的关键分子。由于其在免疫应答中的独特地位,PKCθ已成为自身免疫性疾病和肿瘤免疫治疗领域的重要靶点。
利用PRKCQ基因敲除的HAP1细胞,可以在同基因背景下精准评估该激酶功能缺失对T细胞信号转导的影响,为靶向PKCθ的药物筛选和机制研究提供可靠的细胞模型。
图2. T细胞信号传导的正向调控
● 酪氨酸激酶与神经发育:
CDKL5基因编码一种在神经系统发育中起关键作用的激酶。该基因的致病性变异是CDKL5缺乏症(CDKL5 deficiency disorder,CDD)的病因,患者临床表现以早发性、难治性癫痫和严重发育迟缓为特征。
利用CDKL5基因敲除的HAP1细胞,可在人源细胞背景下研究CDKL5缺失对神经元分化、突触形成和电生理活性的影响。该模型为解析CDD的分子病理机制及筛选潜在治疗药物提供了理想的体外平台。
CDKL5基因编码一种在神经系统发育中起关键作用的激酶。该基因的致病性变异是CDKL5缺乏症(CDKL5 deficiency disorder,CDD)的病因,患者临床表现以早发性、难治性癫痫和严重发育迟缓为特征。
利用CDKL5基因敲除的HAP1细胞,可在人源细胞背景下研究CDKL5缺失对神经元分化、突触形成和电生理活性的影响。该模型为解析CDD的分子病理机制及筛选潜在治疗药物提供了理想的体外平台。
2. 表观遗传与转录调控:揭示癌症与发育的关键开关
组蛋白修饰、DNA甲基化和染色质重塑等表观遗传机制,在基因表达调控中扮演着重要角色。HAP1敲除细胞为研究这些调控因子的功能提供了理想的实验平台。
● 组蛋白修饰与基因转录:
赖氨酸甲基转移酶2C(KMT2C,也称MLL3)是组蛋白H3赖氨酸4(H3K4)甲基转移酶,催化具有转录激活功能的组蛋白修饰标记。大规模肿瘤测序研究表明,KMT2C是多种实体瘤中突变频率最高的表观遗传调控因子之一,其功能缺失可促进肿瘤发生和转移。
利用KMT2C基因敲除HAP1细胞,可在同基因背景下探究KMT2C缺失对染色质状态和基因表达谱的影响,为理解该基因的肿瘤抑制机制提供细胞模型。
赖氨酸甲基转移酶2C(KMT2C,也称MLL3)是组蛋白H3赖氨酸4(H3K4)甲基转移酶,催化具有转录激活功能的组蛋白修饰标记。大规模肿瘤测序研究表明,KMT2C是多种实体瘤中突变频率最高的表观遗传调控因子之一,其功能缺失可促进肿瘤发生和转移。
利用KMT2C基因敲除HAP1细胞,可在同基因背景下探究KMT2C缺失对染色质状态和基因表达谱的影响,为理解该基因的肿瘤抑制机制提供细胞模型。
● 超级增强子与癌基因调控:
BRD2属于BET溴结构域蛋白家族,通过识别乙酰化组蛋白调控基因转录,是超级增强子区域的关键因子。BET抑制剂可通过抑制BRD2等家族成员的功能,下调超级增强子驱动的癌基因(如MYC)表达,是目前抗肿瘤药物研发的热点方向之一。
利用BRD2基因敲除HAP1细胞,可在同基因背景下评估BRD2缺失对MYC等癌基因表达的影响,为BET抑制剂的机制研究和药效评价提供可靠的细胞模型。
BRD2属于BET溴结构域蛋白家族,通过识别乙酰化组蛋白调控基因转录,是超级增强子区域的关键因子。BET抑制剂可通过抑制BRD2等家族成员的功能,下调超级增强子驱动的癌基因(如MYC)表达,是目前抗肿瘤药物研发的热点方向之一。
利用BRD2基因敲除HAP1细胞,可在同基因背景下评估BRD2缺失对MYC等癌基因表达的影响,为BET抑制剂的机制研究和药效评价提供可靠的细胞模型。
3. DNA损伤与修复:解析化疗耐药与遗传病机制
DNA损伤修复通路的缺陷与癌症易感性、化疗耐药及多种遗传病密切相关。HAP1细胞因单倍体特性,在研究DNA损伤修复机制方面具有独特优势。
● 范可尼贫血(FA)通路与DNA交联修复:
FANCE是范可尼贫血核心复合物的关键组分,其功能缺陷导致细胞对DNA交联剂(如丝裂霉素C)的敏感性显著升高。
Moder等人(2017)在HAP1细胞中构建了FANCC功能缺失细胞模型,并结合全基因组CRISPR文库筛选,揭示了BLM解旋酶复合物的缺失可抑制FANCC相关表型,证实了系统性全基因组筛选方法可用于鉴定DNA修复缺陷的遗传抑制相互作用。
FANCE(c.-56C>T)点突变HAP1细胞株正是研究范可尼贫血通路功能和筛选合成致死靶点的有力工具。
FANCE是范可尼贫血核心复合物的关键组分,其功能缺陷导致细胞对DNA交联剂(如丝裂霉素C)的敏感性显著升高。
Moder等人(2017)在HAP1细胞中构建了FANCC功能缺失细胞模型,并结合全基因组CRISPR文库筛选,揭示了BLM解旋酶复合物的缺失可抑制FANCC相关表型,证实了系统性全基因组筛选方法可用于鉴定DNA修复缺陷的遗传抑制相互作用。
FANCE(c.-56C>T)点突变HAP1细胞株正是研究范可尼贫血通路功能和筛选合成致死靶点的有力工具。
● 核苷酸切除修复(NER)与化疗敏感性:
ERCC1是核苷酸切除修复通路的核心因子,与XPF形成异源二聚体,负责切除紫外线或化疗药物(如顺铂)引起的DNA损伤。
早期研究已证实,ERCC1基因缺失会显著降低细胞对顺铂诱导的DNA损伤的修复能力,并增加对铂类药物(顺铂、卡铂、奥沙利铂)的敏感性。
基于ERCC1(c.*590T>C)点突变HAP1细胞株和ERCC4(c.207+11G>A)点突变HAP1细胞株的研究,可在同基因背景下精准量化这些基因缺失后细胞对铂类药物的敏感性变化,为揭示化疗耐药机制和开发增敏策略提供关键细胞模型。
ERCC1是核苷酸切除修复通路的核心因子,与XPF形成异源二聚体,负责切除紫外线或化疗药物(如顺铂)引起的DNA损伤。
早期研究已证实,ERCC1基因缺失会显著降低细胞对顺铂诱导的DNA损伤的修复能力,并增加对铂类药物(顺铂、卡铂、奥沙利铂)的敏感性。
基于ERCC1(c.*590T>C)点突变HAP1细胞株和ERCC4(c.207+11G>A)点突变HAP1细胞株的研究,可在同基因背景下精准量化这些基因缺失后细胞对铂类药物的敏感性变化,为揭示化疗耐药机制和开发增敏策略提供关键细胞模型。
4. 代谢与离子通道:从脂质代谢到神经科学
代谢异常和离子通道功能紊乱是多种代谢性疾病和神经系统疾病的病理基础。HAP1敲除细胞为此类研究提供了精准的工具。
● 脂质代谢与动脉粥样硬化:
低密度脂蛋白受体(LDLR)是清除血液中低密度脂蛋白胆固醇的关键分子,其基因突变是家族性高胆固醇血症(FH)的主要病因。研究表明,LDLR功能缺失导致血浆LDL-C水平显著升高,患者早发动脉粥样硬化性心血管疾病风险增加。
LDLR(c.*52G>A)点突变HAP1细胞株可用于研究LDLR缺失对胆固醇代谢通路的影响。为体外研究家族性高胆固醇血症的分子机制提供了理想的细胞模型。
低密度脂蛋白受体(LDLR)是清除血液中低密度脂蛋白胆固醇的关键分子,其基因突变是家族性高胆固醇血症(FH)的主要病因。研究表明,LDLR功能缺失导致血浆LDL-C水平显著升高,患者早发动脉粥样硬化性心血管疾病风险增加。
LDLR(c.*52G>A)点突变HAP1细胞株可用于研究LDLR缺失对胆固醇代谢通路的影响。为体外研究家族性高胆固醇血症的分子机制提供了理想的细胞模型。
● 离子通道与神经疾病:
钾电压门控通道亚家族C成员1(KCNC1,编码Kv3.1蛋白)在高频放电的抑制性中间神经元中高表达,对神经元的高频放电能力至关重要。
全外显子测序研究已鉴定出KCNC1基因的致病性错义突变与进行性肌阵挛癫痫(EPM7型)密切相关。突变导致Kv3.1通道功能异常,影响GABA能中间神经元的快速放电能力,从而诱发癫痫发作。
KCNC1基因敲除HAP1细胞为研究KCNC1突变如何影响通道功能及神经元电生理特性提供了体外研究平台,有助于解析癫痫的发病机制。
钾电压门控通道亚家族C成员1(KCNC1,编码Kv3.1蛋白)在高频放电的抑制性中间神经元中高表达,对神经元的高频放电能力至关重要。
全外显子测序研究已鉴定出KCNC1基因的致病性错义突变与进行性肌阵挛癫痫(EPM7型)密切相关。突变导致Kv3.1通道功能异常,影响GABA能中间神经元的快速放电能力,从而诱发癫痫发作。
KCNC1基因敲除HAP1细胞为研究KCNC1突变如何影响通道功能及神经元电生理特性提供了体外研究平台,有助于解析癫痫的发病机制。
5. 泛素-蛋白酶体与细胞凋亡:调控蛋白质稳态
泛素-蛋白酶体系统(UPS)调控着细胞内绝大部分蛋白质的降解,其功能异常与神经退行性疾病、癌症等密切相关。HAP1敲除细胞为研究UPS相关基因的功能提供了理想的同基因型模型。
● 免疫蛋白酶体与抗原呈递:
PSMB8编码免疫蛋白酶体的催化亚基β5i,是免疫蛋白酶体的核心组分,在细胞内蛋白质降解、内环境稳定维持和内源性抗原提呈方面发挥重要作用。研究发现,PSMB8基因突变与多种自身炎症性疾病相关,包括Nakajo-Nishimura综合征、CANDLE综合征等。
利用PSMB8基因敲除HAP1细胞,可在同基因背景下研究免疫蛋白酶体功能缺失对抗原加工、细胞因子产生及炎症反应的影响,为解析该类自身炎症性疾病的发病机制和开发靶向治疗策略提供关键的细胞模型。
PSMB8编码免疫蛋白酶体的催化亚基β5i,是免疫蛋白酶体的核心组分,在细胞内蛋白质降解、内环境稳定维持和内源性抗原提呈方面发挥重要作用。研究发现,PSMB8基因突变与多种自身炎症性疾病相关,包括Nakajo-Nishimura综合征、CANDLE综合征等。
利用PSMB8基因敲除HAP1细胞,可在同基因背景下研究免疫蛋白酶体功能缺失对抗原加工、细胞因子产生及炎症反应的影响,为解析该类自身炎症性疾病的发病机制和开发靶向治疗策略提供关键的细胞模型。
● 去泛素化酶与肿瘤抑制:
USP7(又称HAUSP)是一种重要的去泛素化酶,通过去泛素化作用调控p53和MDM2蛋白的稳定性。研究证实,USP7与p53和MDM2形成调控复合物,在维持p53稳态中发挥核心作用。
USP7在多种肿瘤细胞中过度表达,抑制其活性可有效抑制肿瘤发生发展,已成为抗肿瘤药物研发的热门靶点。USP7基因敲除HAP1细胞为筛选靶向USP7的小分子抑制剂、研究USP7-p53/MDM2轴的调控机制提供了可靠的体外模型。
USP7(又称HAUSP)是一种重要的去泛素化酶,通过去泛素化作用调控p53和MDM2蛋白的稳定性。研究证实,USP7与p53和MDM2形成调控复合物,在维持p53稳态中发挥核心作用。
USP7在多种肿瘤细胞中过度表达,抑制其活性可有效抑制肿瘤发生发展,已成为抗肿瘤药物研发的热门靶点。USP7基因敲除HAP1细胞为筛选靶向USP7的小分子抑制剂、研究USP7-p53/MDM2轴的调控机制提供了可靠的体外模型。
03
HAP1——艾迪基因现货基因编辑细胞库
技术核心与优势
● 靶点丰富,覆盖广泛
现货细胞覆盖激酶、表观遗传、DNA损伤修复、代谢、离子通道、神经科学、自噬、去泛素化、肿瘤抑制等多个热门研究方向,满足不同科研需求。
现货细胞覆盖激酶、表观遗传、DNA损伤修复、代谢、离子通道、神经科学、自噬、去泛素化、肿瘤抑制等多个热门研究方向,满足不同科研需求。
● 现货供应,即订即用
细胞库现货充足,下单后快速发货,为您节省2-3个月的自建细胞株时间,让研究更高效。
细胞库现货充足,下单后快速发货,为您节省2-3个月的自建细胞株时间,让研究更高效。
● 全程质控,数据可靠
均通过Sanger测序、支原体检测及STR鉴定,保证细胞身份准确、无污染,实验结果可重复。
均通过Sanger测序、支原体检测及STR鉴定,保证细胞身份准确、无污染,实验结果可重复。
● 产品线完整,一应俱全
提供KO敲除细胞、PE点突变细胞、OE稳转株(Cas9、GFP、荧光素酶)及WT野生型对照细胞,满足从基因敲除到功能验证的全流程需求。
提供KO敲除细胞、PE点突变细胞、OE稳转株(Cas9、GFP、荧光素酶)及WT野生型对照细胞,满足从基因敲除到功能验证的全流程需求。
完整清单包含数百种基因靶点,涵盖激酶、表观遗传、DNA修复、代谢、离子通道等多个研究方向。更多产品及定制服务,请联系艾迪基因客服。
04
结语
从全基因组CRISPR筛选到疾病相关点突变的功能解析,从线粒体多组学研究到药物靶点的验证——HAP1基因编辑细胞凭借其独特的单倍体优势,已成为全球科研人员加速科学发现的核心工具之一。Rensvold等人在2022年发表的MITOMICS计划,更是将HAP1细胞的应用推向了系统性、多组学、高通量的新高度。
艾迪基因现货HAP1细胞库,覆盖数百个基因靶点,每株细胞均经过严格验证,即订即发,助您的研究快人一步。
参考文献
[1]. Osório, J., et al. (2015). Functional genomics: The genetic essence of human cells. Nature Reviews Genetics, 16(12), 683.
[2]. Rensvold, J. W., et al. (2022). Defining mitochondrial protein functions through deep multiomic profiling. Nature, 606(7913), 382–388.
[3]. Smits, A. H., et al. (2019). Biological plasticity rescues target activity in CRISPR knock outs. Nature Methods, 16(11), 1087–1093.
[4]. Kalaitzidis, D., et al. (2012). mTOR complex 1 plays critical roles in hematopoiesis and Pten-loss-evoked leukemogenesis. Cell Stem Cell, 11(3), 429–439.
[5]. Moder, M., et al. (2017). Parallel genome-wide screens identify synthetic viable interactions between the BLM helicase complex and Fanconi anemia. Nature Communications, 8(1), 1238.
[6]. Leonard, H., et al. (2022). CDKL5 deficiency disorder: clinical features, diagnosis, and management. The Lancet Neurology, 21(6), 563–576.
联系我们
18102225074(微信同号)
market@edgene.cn
标签
Share








